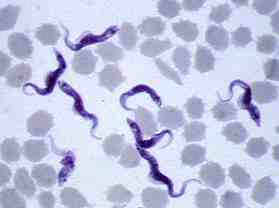
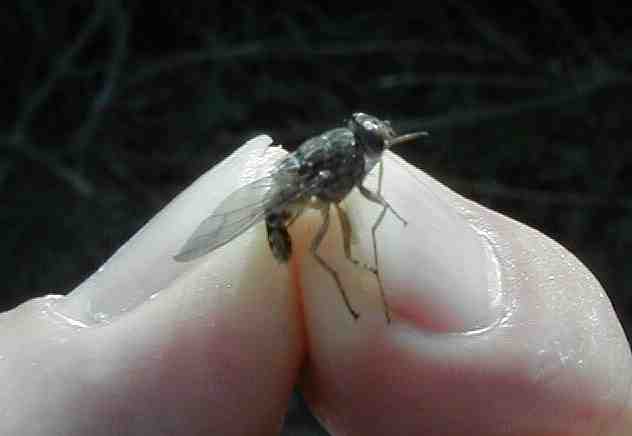
何種類かのトリパノソーマ原虫によって引き起こされるアフリカ・トリパノソーマ症は、20数種の吸血性のツェツェバエ(写真)によって極めて効率よく媒介されている。原虫が昆虫体内でも増えるため、ツェツェバエ
Tsetse fly: Glossina spp. は「ベクター」と呼ばれる。原虫はヒトや動物の流血中(写真)で増殖し、トリパノソーマ症
Trypanosomosis(かつては Trypaosomiasis と呼ばれていた) を起こし、死に至る。人畜(獣)共通伝染病のひとつであり、ヒトではガンビア・トリパノソーマ
Trpanosoma brucei gambiense による西アフリカには慢性型、東アフリカにはローデシア・トリパノソーマ
Trypanosoma brucei rhodesiense 急性型の眠り病が知られている。動物(野生生物、家畜)では慢性経過をとることが多く(そうでないと、原虫は絶滅してしまう)、著明な貧血、泌乳量・体重の減少、流産そして死に至ることが問題となっている。
現在、サブサハラ・アフリカの37の国々において、ウシだけでも6000万頭以上が感染の危機に曝されている。年間の直接及び間接的な経済的損失だけでも、50億米ドル以上と推定されている。
今世紀の初め、ビクトリア湖周辺部、今日のウガンダでは多数の患者が発生し、細菌学の父ロベルト・コッホも原因究明のため当地を訪れている。彼が撮影したといわれる下の写真は当時の悲惨な状況を今に伝えている。
一方、北アフリカから中近東・南アジア・東南アジアにかけては、アブが機械的に媒介(単なる「キャリヤー」=運び屋)する T. evansi が引き起こすトリパノソーマ症が散発し、問題となっている。また、南米には吸血性のカメムシであるサシガメの媒介する T. cruzi が引き起こすシャーガス病が知られているが、この原虫は細胞内寄生をするという点で、上記のトリパノソーマ症と異なる。
また、トリパノソーマ原虫はトランス・ローケーションやトランス・スプライシングにより変異表面糖タンパク質(VSG)を次々に脱ぎ替えたり、キネトプラスト(特殊に変化したミトコンドリアの部分)では転写後のRNAが更に編集されること(RNA editing)で、長年にわたって分子生物学者の興味の対象となってきている。
私は、ここで今までの形態学的、生態学的比較からなされた原虫の分類が、いかに不十分で実状に合っていないかを具体的な事例としてトリパノソーマ原虫の分類の問題点を指摘し、遺伝子の比較も考慮した分類がなされるべきかについても述べたい。

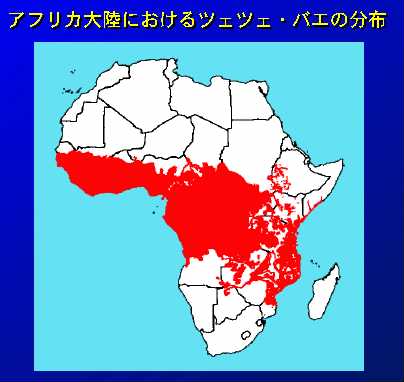
19世紀、『暗黒アフリカ大陸の探検・発見』で有名な欧米の探検家たちは、数百頭のウシを引き連れキャラバンを組んでアフリカ大陸を縦横した。と同時に、熱帯地方の限られたツェツェバエの生息地をいっきに広めたとされる。その結果、今日では、砂漠以外の赤道を挟む広大なアフリカの農業可能地域には、ツェツェバエが広く分布しツェツェ・ベルトを形成している。そして、その大部分は同時にトリパノソーマ症で汚染されている。ツェツェバエは一生のうち10匹程度しか子孫をつくらないにも関わらず、卵胎生のため、産まれた幼虫(蛆)はすぐに地中に潜り、僅か2時間ほどで黒化して蛹になる。こうして、アリや小鳥から捕食される危険の多い幼虫の時期をほとんどなくすことで、極めて効率よく繁殖している。

家畜(ウシ、ヒツジ、ヤギ、ウマ、ラクダなど)が感染した場合、アフリカでは「ナガナ」、アフリカ以外では「スーラ」と呼ばれる貧血・痩削が主徴の消耗性の感染を呈し、最後は死に至り経済価値を著しく損ねる。なかでも、アフリカでは、コンゴ・トリパノソーマ
Trypanosoma congolense による被害が、最も深刻である。

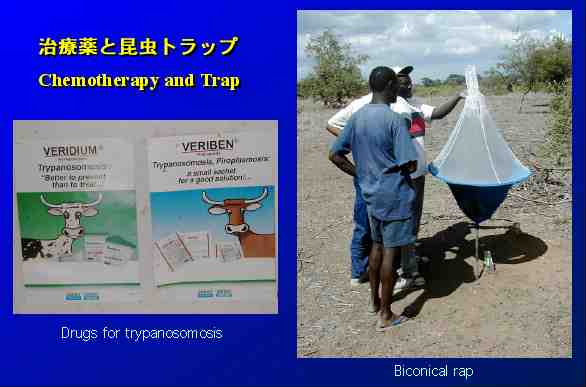
治療には植民地時代の35年以上も前に開発されたままの抗トリパノソーマ薬が、いまだに使われているが、近年マラリアと同様に原虫が薬剤耐性を獲得しつつある。重篤なヒトの感染のケースでは、今も毒性が強くて危険な砒素剤も治療に使われている。予防法は無きに等しく、トラップや環境への悪影響が懸念される殺虫剤でツェツェバエをある程度コントロールするくらいで、新しい化学療法剤と有効なワクチンの開発が望まれている。
1992年から1995年の3年間、私はJICA(国際協力事業団、ジャイカ)の派遣専門家として、ケニアの首都ナイロビにある国際農業研究機関のひとつ、国際動物病研究所(ILRAD:
International Laboratory for Research on Animal Diseases、イルラッド)で研究する機会を得た。研究所は1995年1月に別の国際機関ILCAと統合して、国際畜産研究所(ILRI:
International Livestock Research Institute、イルリ)と名前を変えたが、過去20年にわたりアフリカにおけるトリパノソーマ及びタイレリア原虫感染症研究を継承している。
ILRADでの研究の一つにリボソームRNA遺伝子の解析がある。目的は幾つかの種類のトリパノソーマのリボソーム遺伝子の配列を比較すること、及び転写開始点であるリボソーム遺伝子プロモーター部位の解析であった。ことに、混乱しているコンゴ・トリパノソーマ
T. congolense の分類を明確にすることであった。
遺伝子レベルで原虫(原生動物)を比較・進化を論ずることは、リボソームRNA遺伝子を用いて既になされている。例えば、テトラヒメナ、マラリアとそれが属する Apicomplexa 、コクシジウムの論文がある。私は、その後の研究成果も交え、このページでトリパノソーマ原虫のリボソームRNA遺伝子の解析を通じて、新たな分類の試みについて述べる。
尚、この研究は Phelix A.O.Majiwa博士との共同研究であり、関心のある方は原文をご覧いただきたい。

リボソームは細胞内のタンパク合成器官で、ウイルスを除くすべての生物が持っている。リボソーム粒子は大小2つの粒子から成り、それらは幾つかのRNA鎖(種によってサイズが異なるが、28S,18S,5.8S,5S
RNAがある)と数十種類のリボソーム・タンパク質とから構成されている。
これはカエルのリボソームRNA遺伝子DNAが盛んにRNAに転写されている様子を示す電子顕微鏡像で、その形状からランプ・ブラシとか、クリスマス・ツリーなどと表現されている。多くの生物ではタンパク質を効率的に合成するために、たくさんのリボソームが必要なことから、リボソーム遺伝子がタンデム(同じ方向で直列)にリピートしている。これらの一つ一つのリボソーム遺伝子の配列は極めてよく似ており、協調進化
Concerted evolution の代表例として分子生物学の教科書に載っている。
トリパノソーマ原虫も、数は確定されていないが、ゲノムライブラリーからのクローニングの効率の良さから考えて、数十以上あると思われる。こうして新生されたリボソームRNA前駆体は、その後直ちにRNA分解酵素によって不必要な部分=スペーサーを切り取られた後、数十のリボソーム・タンパクと結合して80Sラージ・サブユニットと40Sスモール・サブユニットを形成する。
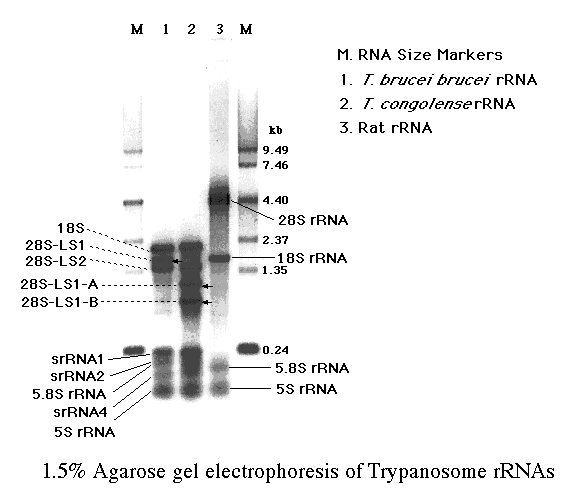
トリパノソーマから全RNAを抽出して、寒天ゲル電気泳動で分析した。全RNAの95%程度がリボソームRNAで、残り数%がmRNAやtRNAなどである。トリパノソーマはラットなどの高等生物と違って、4,500ヌクレオチド程の28S
rRNA に相当するバンドが無いことに注意してもらいたい。
更に、T. brucei では明確な28S-LA1のバンドが T. congolense では極めて薄く、LS1-AとLS1-Bの2つのバンドが新たに見られる。この現象については最後に述べる。

この実験で使ったトリパノソーマのリストを上に掲げた。T. congolense のうちサバンナ・タイプは最も広範囲に分布しており、分離地域・宿主・分離年の異なる5株を調べた。T.
congolense の変種とされるツァボTsavo・タイプ、ケニアの海岸地方のキリフィーKilifi・タイプ、西アフリカ・河川森林(WARF)・タイプも加えた。T.
simiae はブタ類のトリパノソーマで、名前から類推されるサルとは直接関係がない。最初にサルを使って分離されたのが命名の由来で、誤解を招くこの種名は一日も早く改正されるべきだ。
以上が、アフリカのトリパノソーマであるが、最後の T. evansi はタイの首都バンコクに近いパトンタニ Pathum Thani にあるウシの人工受精センターでの小規模な流行から、マウスで分離された株である。後に、台湾の黄牛(?)からの分離株からもデータを得た(: D89527)。
ちなみに、各遺伝子の Accession number を以下にリンクしておく。
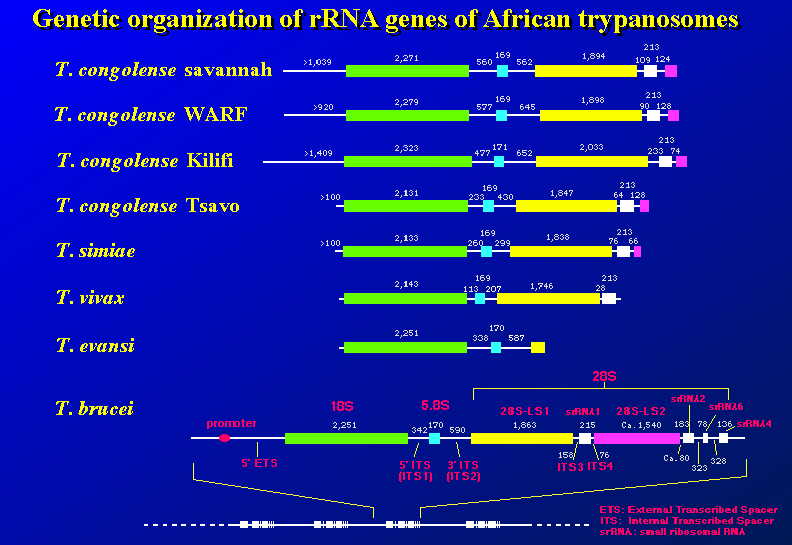
この実験では、原虫のゲノムDNAからファージDNAライブラリーを作り、リボソーム領域を含むクローンを選抜し配列決定がなされた。得られた遺伝子地図とそのアクセッション・ナンバー(GenBank)を示した。T.
brucei brucei のデータは GenBank のデーター・ベースから引用した。それ以外は全てこの研究で得られた。
リボソーム遺伝子DNAからリボソームRNAへの転写は、RNAポリメレースIによって18S RNAの上流数Kbにあるプロモーター領域から始まり、28S領域の下流で終了する10Kベース近くの長い前駆体RNAが一旦合成される。この分子は直ちにRNA分解酵素により特異的切断を受け、18Sと5.8Sと28SのRNA鎖ができる。5S RNAは染色体の別の部位からRNAポリメレースIIIによって、転写される。
トリパノソーマでは、28S RNAが比較的大きな2つのフラグメントと、4つの小さなフラグメントに切断されている。18S RNAの配列は分子進化の指標として、種々の生物種で解析されてきている。今回我々は18S rDNAと28S-LS1 rDNAの両方の配列を比較した。
また、スペーサー部分の長さは種によって異なるために、大まかな分類の指標になりうる。例えば18Sと5.8Sとの間の5'ITSは、PCRで、わずかの長さの違いを検出しやすい部位である。
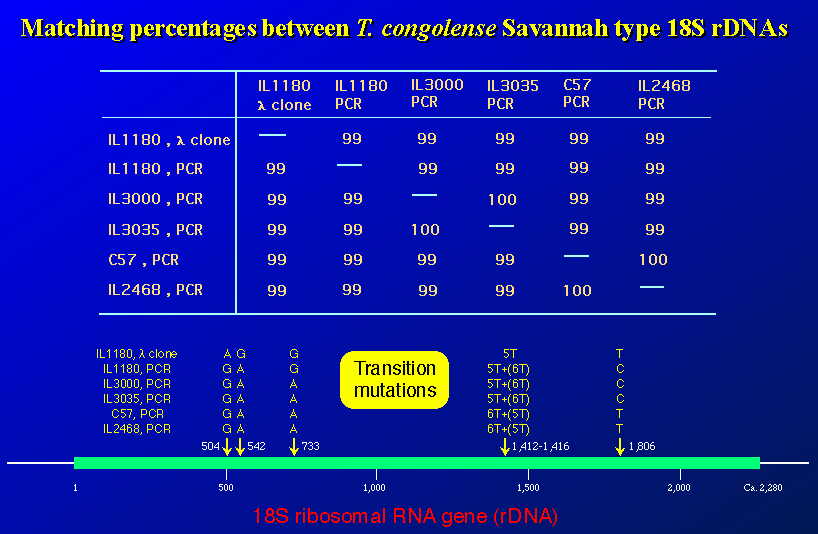
アフリカ各地からの T. congolense サバンナ・タイプ分離株間の18S RNA遺伝子領域での違いを示した。PCR産物はプラスミドへサブクローニングすることなしに、直接法で塩基配列を決定した。全長2,300ベース足らずの18S
rDNA遺伝子の僅かに5カ所に変異が認められた。うち4つはトランジッション・ミューテーション(プリン、ピリミジンどうし)であり、残りの一つはポジション・ナンバー1412から始まるチミンの数が5つが多いか、6つが多いかの違いであった。IL1180では同じ株由来の配列がラムダ・ファージ由来のクローンとPCR産物からの直接法での配列とで違いが見られた。
得られた18S rDNAの配列を相互に比較してみた。90%以上の場合を太字で示した。リーシュマニア(ヒトや動物の疾病を起こす)とクライシディア(昆虫の消化管内で自由生活をしている)は同じトリパノソーマ科に属する生物で、データーベースから引用した。サバンナ・タイプとWARFタイプが比較的近く、その次にKilifiタイプが近い。どちらかと言うTsavoタイプは
T. simiae に近い。以外なのは、リーシュマニアとクライシディアが遺伝的に近い関係にあることが分かる。
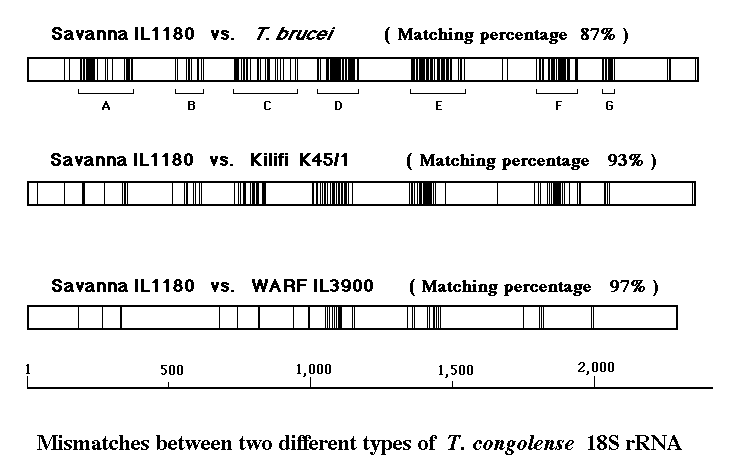
これらの違いを遺伝子上にプロットしてみると、18S遺伝子の全体にバラバラと存在するのではなく、幾つかの「かたまり」=クラスター(A~G)を形成していることが分かる。
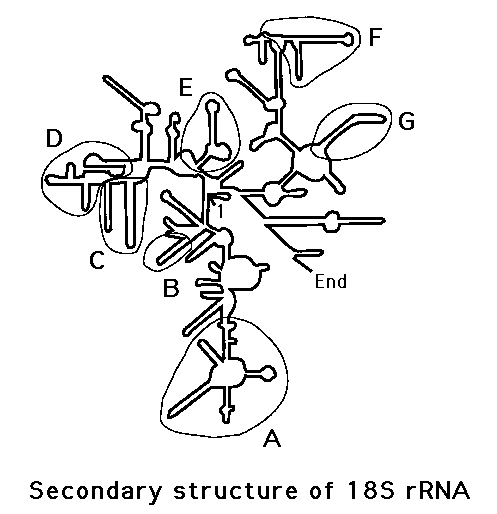
リボソームRNAは1本鎖の故、相補部分が水素結合を成して、高次構造をとることが知られているが、これらのクラスターは「枝葉」=ループ部分に当たり、よく保存されているところは、「幹」=ステム部分に当たることが分かった。

同じ株が持つ複数のリボソーム遺伝子どうしの塩基配列は極めてよく保存されているが、リボソーム遺伝子間にあって、RNA分解酵素で切り取られるスペーサー部分には多くの変異が認められた。
ここでは、18Sの前約450ベースをラムダ・ファージ・ライブラリー由来と PCR産物からサブクローニングされたクローンの配列を比較してみた。図のようにこの部位はホモロジーがあるものの、塩基置換や欠損が数多く存在する。他の部分のスペーサーにもこの様な現象が観察された。
これらの結果は、『自然淘汰にかかりにくい(中立=有害でない)と思われるほど分子レベルでは進化速度が大きい』と言う、木村資生の分子進化論である「中立説」(木村資生著:『分子進化を考える』岩波新書)を支持するものである。
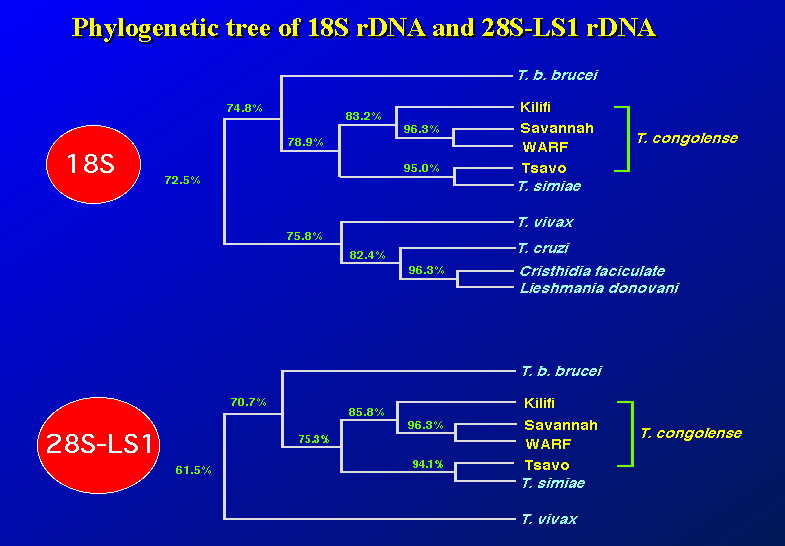
18Sと28S-LS1の領域をそれぞれコンピューター処理し、系統樹を描かせてみた。どちらの系統樹も僅かな数値の違いがあるのみで、殆ど同じ結果を得た。また、タイのウシ由来と台湾のスイギュウ由来の
T. evansi は、T. brucei brucei との間で、18S RNA遺伝子に、僅か2ベースの違い(ポジション243とポジション345)があるだけだったので、ここでは表示していない。このことは、
T. evansi が T. brucei からつい最近(数1000年前?)派生した変異株 variant
であることを意味している(詳しくは次の「アフリカを飛び出したアフリカ・トリパノソーマ」を参照)。私は、更に詳しい遺伝子解析を現在(2000年3月)行っており、T.
evansi の起源の解明やワクチン開発を目指している。
サバンナ・タイプに一番近いのはWARFタイプで、Kilifiタイプはそれからやや離れている。Tsavoタイプは T. simiae に近く、上記の三者と離れている。
T. vivax は T. brucei や T. congolense のグループよりも、T. cruzi に近いのも意外である。
面白いのは、18S rDNAの配列を見ると、 Cristhidia faciculate と Leishmania donovani とがかなり近いことである。これら分類学上離れている原虫が遺伝子レベルで見ると近い。このことは、これらの進化の歴史の上から見れば、興味ある事実である。

トリパノソーマを含め、原虫(単細胞生物)の分類は、形態的特徴の他、媒介性、宿主域など、もっぱら表現形を基にしてなされてきた。Trypanosoma
属は、唾液が感染に関連する Salivaria 区(Section)と昆虫の排泄物を通して伝搬されるグループ Stercoraria
区(Section)に大別される。
Salivaria 区には Trypanozoon 、Nannomonas、Duttnella、Pycnomonas 亜属が属する。 Nannomonas 亜属(小さい原虫の意)には T. congolense と T. simiae の2種が属する。 最も重要な T. congolense はサバンナ・タイプが殆どであるが、亜種(あるいは変種)として他にKilifi、WARF、Tsavo タイプが形態的特徴から入れられている。
Trypanozoon 亜属には T. evansi、T. brucei、T. equiperdum が属す。T. brucei は更に T. brucei brucei、T. brucei gambiense、T. brucei rhodesiense の3亜種に分けられている。
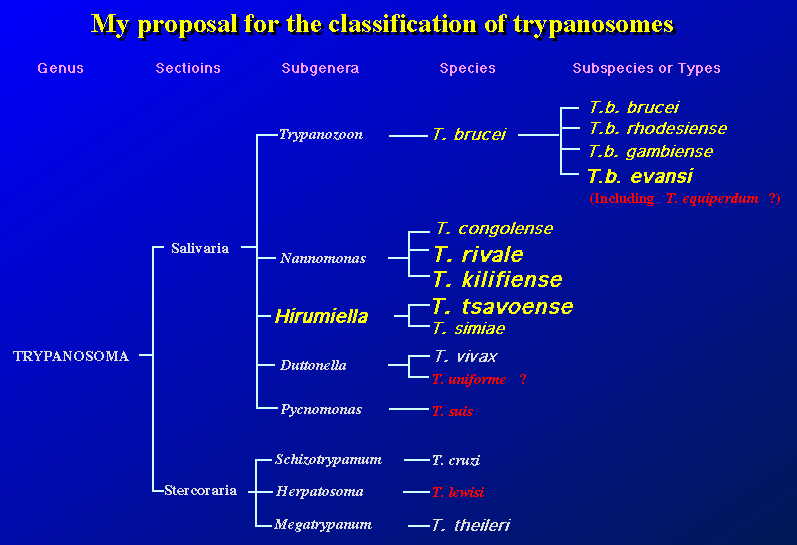
私の提案している分類は、従来の形態的分類に加えて、遺伝子レベルでの違いを加味したものである。従来の分類との大きな違いは T.
congolense のサバンナ・タイプを独立させ、WARFタイプを T. rivale、Kilifiタイプを
T. kilifiense とし、Tsavo タイプをより近い T. simiae と一緒にし、新たな
Subgenera(亜属)として Hirumiella の新設を提案したい。
遺伝レベルで見れば、極めて近い T. evansi を T. brucei に編入し、T. brusei の亜種とする。この種 T. brucei は、現在、分布と宿主と生活環を変えつつ、新たな種への分化を模索しつつあるようだ。
プライマー伸展法 Primer extention により、T. congolense の28S-LS RNA分子の切断点を調べてみた。リボソームRNAと適当なアンチセンスのDNAプライマーを混ぜ、逆転写酵素を反応させる。一方、同じプライマーを用いて、サンガー法でリボソーム遺伝子DNAの塩基配列を決める。両者を同一ゲルで電気泳動した後、オートラジオグラフィーをした。
念のため、18Sと28Sの5'の切断点が正確に行われているかを確かめた。この実験の結果、 T. congolense の28S-LS RNA分子の切断点は2つあることが示唆された。
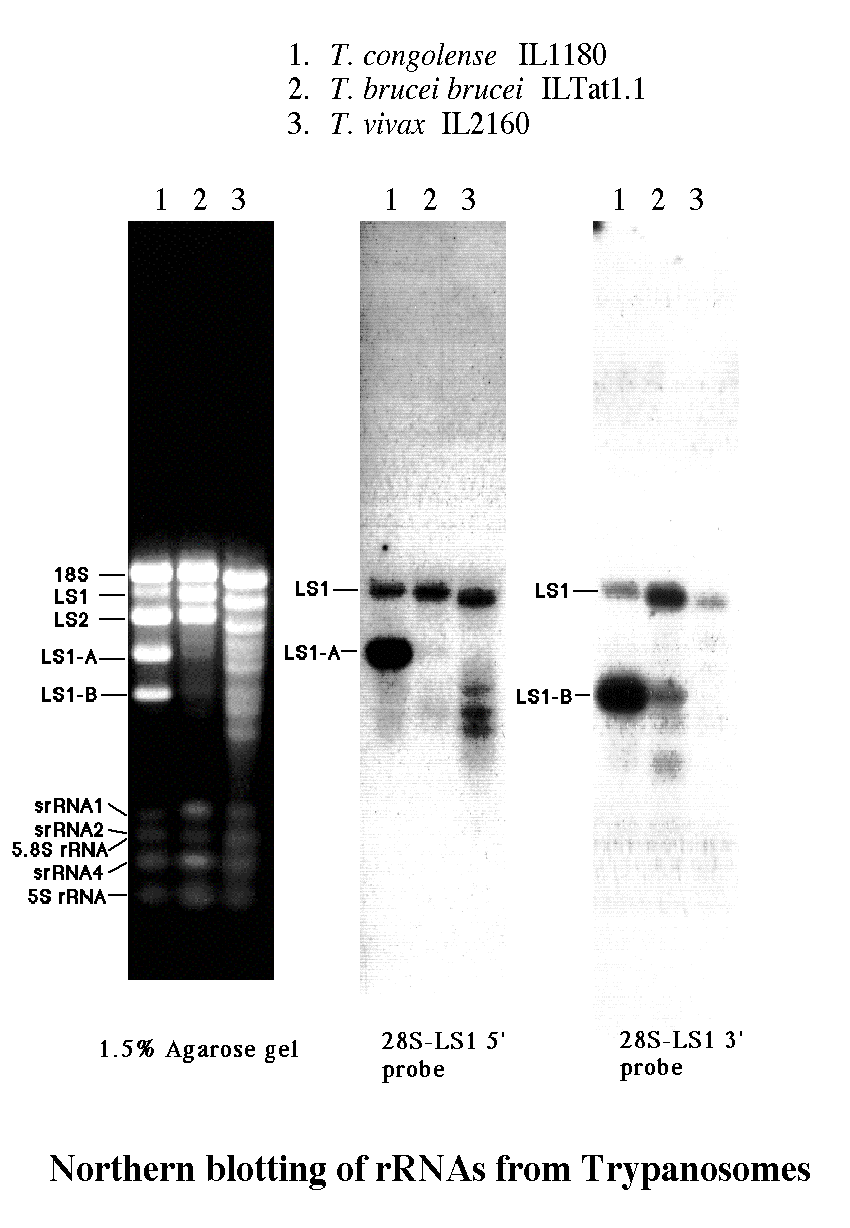
T. congolense、T. brucei、T. vivax から調製した全RNAの電気泳動像で、T.
congolense では28S-LS1のバンドが薄くなっており、28S-LS1-A(1,200ベース)と28S-LS1-B(700ベース)の2つのバンドが出現している。T.
brucei にも T. vivax にもその辺りにごく薄いバンドが見られた。
Primer extention test で、T. congolense の28S-LS1の切断点が2カ所あることが示唆されたので、どちらが主要な切断点かを調べるために、28S-LS1分子の5'側400ベースと3'側300ベースのプローブをそれぞれ当ててみた。するとLS1-Aバンドは5'側プローブとよく反応し、LS1-Bバンドは3'側プローブとよく反応した。T. brucei では、3'側プローブを用いた時、LS1-Bに相当する薄いバンドが見られた。
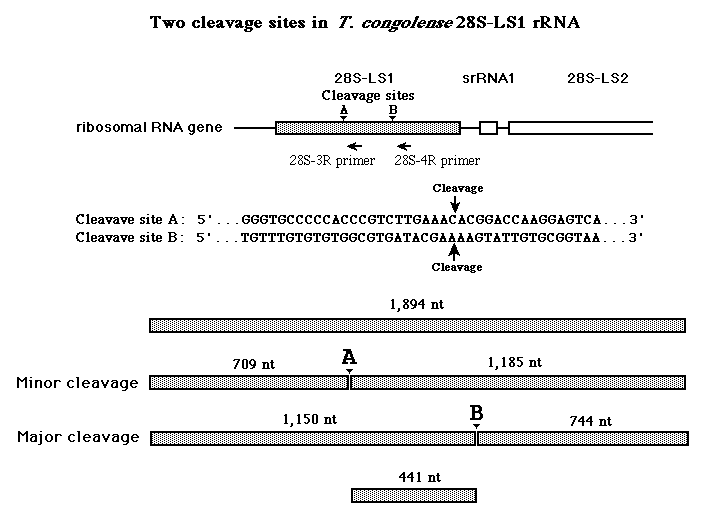
以上をまとめると、T. congolense では上の図のように主要な切断点はBであることが示唆された。T.
brucei では T. congolense の切断点Aにあたる部分と相同な塩基配列を保持しており、その部位で
T. brucei でもごく一部の28S-LS1が切断を受けているようだ。
また、上流のプロモーター部位の配列も調べたが、ここでは触れない。


これらの研究を進める際、蛭海先生ご夫妻(元 ILRAD・帯広畜産大学)と良き共同研究者 Phelix Majiwa 博士の絶大なる協力があったことに感謝し、ここに記しておく。
最後に、この文章は私一個人の意見であって、ILRAD(現ILRI)やJICA(国際協力事業団)の方針や見解を表したものではないを断っておく。それ故、内容に関して、私
浦川豊彦が全責任を負うものである。
![]()